
搜索结果: 1-15 共查到“基础医学 恶性疟原虫”相关记录110条 . 查询时间(0.337 秒)
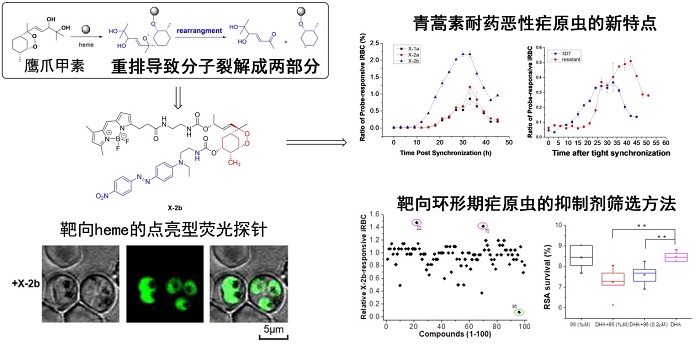
2022年5月18日,中国医学科学院药物研究所天然药物活性物质与功能国家重点实验室庾石山团队、张崇敬团队和中国医学科学院基础医学研究所王恒团队在国际著名期刊Signal Transduction and Targeted Therapy杂志发表了题为“A heme-activatable probe and its application in the high-throughput screen...

2022年3月14日,中南大学医学寄生虫学系教师尚晓敏与同济大学医学院张青锋课题组合作,在国际知名权威学术期刊《Nucleic Acids Research》 (IF=16.971)杂志在线发表题为 “Genome-wide landscape of ApiAP2 transcription factors reveals a heterochromatin-associated regulato...
我国科学家研发出针对恶性疟原虫的新型遗传操作工具
科学家 恶性疟原虫 新型遗传操作工具
2019/1/7
疟疾与艾滋病、结核病一起被列为全球三大传染性疾病。疟原虫是引起疟疾的病原体,其中恶性疟原虫的感染致死率最高。分子水平的遗传操作是研究恶性疟原虫致病机理以及抗药机制的重要手段。然而,在恶性疟原虫中通过同源重组方法进行基因编辑的效率极低,而且恶性疟原虫缺乏可运用RNAi方法来进行基因编辑的关键原件,因此对恶性疟原虫的研究急需发展一种高效简便的遗传操作工具。

2018年12月24日,国际学术期刊《美国国家科学院院刊》(PNAS)在线发表了中国科学院上海巴斯德研究所江陆斌研究组题为Epigenetic editing by CRISPR/dCas9 in Plasmodium falciparum 的最新研究成果。疟疾与艾滋病、结核病一起被列为全球三大传染性疾病。疟原虫是引起疟疾的真核病原微生物,其中恶性疟原虫的感染致死率最高。分子水平的遗传操作是研究恶...
基于恶性疟原虫富含组氨酸蛋白2抗原诊断方法的研究进展
恶性疟原虫 富含组氨酸蛋白 快速诊断试剂
2015/2/2
恶性疟原虫是引起人类疟疾的主要病原体之一,采用基于恶性疟原虫富含组氨酸蛋白2(Plasmodium falciparum histidine-rich-protein 2,PfHRP2)抗原的快速诊断试剂(rapid diagnostic test, RDT)进行诊断是诊断恶性疟原虫感染的特异性方法,但表达该抗原的pfhrp2基因呈现明显的多态性,甚至表达缺失,导致试剂的灵敏度降低,其临床实用性受...
恶性疟原虫组氨酸富集蛋白Ⅱ、Ⅲ的序列多态性分析
恶性疟原虫 富组氨酸蛋白Ⅱ 富组氨酸蛋白Ⅲ 分型
2014/11/3
分析恶性疟原虫富组氨酸蛋白Ⅱ/Ⅲ(PfHRPⅡ/Ⅲ)的序列多态性。 方法 采集云南疟疾流行区恶性疟患者血样20份,血涂片后镜检,并进行疟疾快速诊断试剂检测(RDTs),同时提取各血样中恶性疟原虫基因组DNA,进行PCR扩增Pfhrp2和Pfhrp3核酸片段并测序。使用Bioedit软件对Pfhrp2和Pfhrp3基因序列进行比对,使用TRANSEQ软件推导出其编码的氨基酸序列。 结果 20份...
恶性疟原虫裂殖子表膜蛋白MSPDBL2-DBL2结构域的克隆表达和抗原性分析
恶性疟原虫 裂殖子 原核表达 抗原性
2014/11/3
克隆、表达恶性疟原虫(Plasmodium falciparum)疫苗候选分子-裂殖子表膜蛋白MSPDBL2 (PF10_0355)的DBL2结构域,并分析其抗原性。 方法 PCR扩增实验室培养的恶性疟原虫标准株3D7的基因组DNA,采用无缝克隆技术快速连接目的片段和质粒载体,构建重组表达质粒pET28a-DBL2,转化至大肠埃希菌BL2l(DE3)中,异丙基-β-D-硫代半乳糖苷(IPTG)诱...
恶性疟原虫顶端膜抗原1基因多态性分析
恶性疟原虫 顶端膜抗原1 基因多态性
2014/11/3
研究不同地理分离株恶性疟原虫裂殖子顶端膜抗原1(PfAMA-1)基因的多态性。 方法 采集2006-2012年福建省23例输入性恶性疟患者的血样,以血样中的疟原虫DNA为模板,巢式PCR扩增AMA-1基因片段,利用生物软件进行序列比对分析。 结果 23份血样均扩增出目的条带(约505 bp),发现32个多态位点,共计18个单倍型,其中8个为新报道序列。来自非洲的恶性疟原虫分离株的AMA-1基因...
输入性恶性疟原虫Pf60.1基因多态性初步研究
输入性病例 恶性疟原虫 Pf60.1 基因多态性
2014/9/25
采用PCR方法扩增武汉市2010-2013年经镜检和巢式PCR确诊的101份国外输入性恶性疟病例血样的Pf60.1基因,研究Pf60.1基因多态性。PCR结果显示,共92份血样扩增出3类基因片段,其中52份扩增出313 bp片段,占56.5%;34份扩增出340 bp片段,占37.0%;6份扩增出313 bp和340 bp混合型片段,占6.5%。83份自非洲地区输入病例血样中,46份扩增出313 ...
恶性疟原虫信号肽肽酶-绿色荧光蛋白突变株的建立及其在疟原虫体内的表达分析
信号肽肽酶 绿色荧光蛋白 转染
2014/9/25
构建恶性疟原虫(Plasmodium falciparum)信号肽肽酶(PfSPP)基因转染载体,筛选可在体内表达疟原虫信号肽肽酶-绿色荧光蛋白(PfSPP-GFP)的疟原虫。 方法 提取Trager-Jensen法培养的恶性疟原虫3D7株基因组DNA,PCR扩增PfSPP C端不含终止密码子的883 bp基因片段,克隆构建重组转染载体pSPPcGT。重组载体经PCR和双酶切鉴定后送测序。电转化...
恶性疟原虫ApiAP2蛋白与var基因内含子序列相互作用的体内鉴定
恶性疟原虫 ApiAP2 var基因 内含子调控蛋白
2014/3/24
鉴定恶性疟原虫ApiAP2蛋白家族成员PF3D7_1107800与var基因内含子保守序列(iNPE)存在体内相互结合作用。 方法 提取恶性疟原虫标准株3D7基因组DNA,PCR扩增PF3D7_1107800基因5′端片段。扩增产物经双酶切后连接入原核表达载体pGex-4T-1,重组质粒转化至大肠埃希菌(E. coli)BL21(DE3),异丙基-β-D-硫代半乳糖苷(IPTG)诱导表达。十二...
短寡核苷酸链高效转染体外培养恶性疟原虫的研究
恶性疟原虫 Entranster-R转染 寡核苷酸链
2014/2/18
5%山梨醇连续2次同步化恶性疟原虫培养物(8 h窗口),培养16 h后,直接孵育组(A组)将50 μl含寡核苷酸培养基与450 μl恶性疟原虫培养物(5%虫血率,1%血压积)混合孵育,Entranster-R试剂转染组(B组)将50 μl转染复合物(含寡核苷酸链和转染试剂)与450 μl恶性疟原虫培养物混合孵育,培养5 h后重悬,分别取出250 μl,1 500×g离心3 min,收集沉淀,进行荧...
短寡核苷酸链高效转染体外培养恶性疟原虫的研究
恶性疟原虫 Entranster-R转染 寡核苷酸链
2014/2/18
5%山梨醇连续2次同步化恶性疟原虫培养物(8 h窗口),培养16 h后,直接孵育组(A组)将50 μl含寡核苷酸培养基与450 μl恶性疟原虫培养物(5%虫血率,1%血压积)混合孵育,Entranster-R试剂转染组(B组)将50 μl转染复合物(含寡核苷酸链和转染试剂)与450 μl恶性疟原虫培养物混合孵育,培养5 h后重悬,分别取出250 μl,1 500×g离心3 min,收集沉淀,进行荧...
恶性疟原虫含s48/45结构域蛋白家族的研究进展
疟疾 恶性疟原虫 s48/45结构域 半胱氨酸
2013/11/11
s48/45结构域表现为β三明治结构,一般含有6-半胱氨酸(6-Cys)。含s48/45结构域的蛋白存在于疟原虫发育阶段的各个时期,而且在虫体入侵宿主细胞的过程中发挥重要作用。根据蛋白分子的特征和功能,发现s48/45蛋白家族可作为恶性疟原虫不同时期(如蚊期、红细胞外期和红细胞内期)的疫苗候选分子。本文主要阐述了恶性疟原虫含s48/45结构域蛋白家族的研究进展。
2011年6月至12月在云南中缅边境,采用双盲法对475例48 h内有发热史的门诊患者进行疟原虫检测。结果表明,使用SDBIOLINE恶性疟原虫/间日疟原虫抗原快速检测试剂盒共检出疟原虫阳性202例,其中恶性疟原虫98例,间日疟原虫104例。镜检结果显示,疟原虫阳性共206例,其中恶性疟99例,间日疟107例。以镜检法为金标准,SDBIOLINE试剂盒检测疟原虫总体敏感性为98.1%(202/20...